
Procariotas marinos - Marine prokaryotes
.jpg)
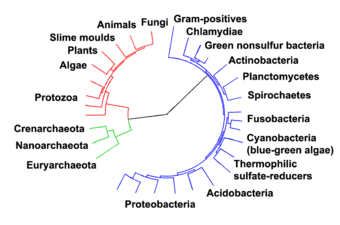
Los procariotas marinos son bacterias marinas y arqueas marinas . Se definen por su hábitat como procariotas que viven en ambientes marinos , es decir, en el agua salada de mares u océanos o en el agua salobre de los estuarios costeros . Todas las formas de vida celular se pueden dividir en procariotas y eucariotas. Los eucariotas son organismos cuyas células tienen un núcleo encerrado dentro de membranas , mientras que los procariotas son organismos que no tienen un núcleo encerrado dentro de una membrana. El sistema de tres dominios para clasificar la vida agrega otra división: los procariotas se dividen en dos dominios de la vida, las bacterias microscópicas y las arqueas microscópicas, mientras que todo lo demás, los eucariotas, se convierte en el tercer dominio.
Los procariotas juegan un papel importante en los ecosistemas como descomponedores que reciclan nutrientes. Algunos procariotas son patógenos y causan enfermedades e incluso la muerte en plantas y animales. Los procariotas marinos son responsables de niveles significativos de la fotosíntesis que ocurre en el océano, así como del ciclo significativo del carbono y otros nutrientes .
Los procariotas viven en toda la biosfera . En 2018 se estimó que la biomasa total de todos los procariotas del planeta era equivalente a 77 mil millones de toneladas de carbono (77 Gt C). Se compone de 7 Gt C para arqueas y 70 Gt C para bacterias. Estas cifras se pueden contrastar con la estimación de la biomasa total de los animales en el planeta, que es de aproximadamente 2 Gt C, y la biomasa total de los humanos, que es de 0.06 Gt C. Esto significa que las arqueas colectivamente tienen más de 100 veces la biomasa colectiva de humanos y bacterias más de 1000 veces.
No hay evidencia clara de vida en la Tierra durante los primeros 600 millones de años de su existencia. Cuando llegó la vida, estuvo dominada durante 3.200 millones de años por los procariotas marinos. La vida más compleja, en forma de eucariotas corona , no apareció hasta la explosión del Cámbrico hace apenas 500 millones de años.
Evolución
|
−4500 -
-
−4000 -
-
−3500 -
-
−3000 -
-
−2500 -
-
−2000 -
-
−1500 -
-
−1000 -
-
−500 -
-
0 -
|
procariotas marinos |
|
||||||||||||||||||||||||
(hace millones de años )
Los procariotas marinos han sido la forma de vida dominante durante la mayor parte de la historia de la Tierra, quizás porque el agua los protegió de la radiación ionizante.
| ||||||||||||||||||||||||||
La Tierra tiene unos 4.540 millones de años. La evidencia indiscutible más antigua de vida en la Tierra data de hace al menos 3.500 millones de años, durante la Era Eoarqueana, después de que una corteza geológica comenzara a solidificarse después del Eón Hadeano fundido anterior . Se han encontrado fósiles de esteras microbianas en areniscas de 348 mil millones de años en Australia Occidental .

Las especies pasadas también han dejado registros de su historia evolutiva. Los fósiles, junto con la anatomía comparada de los organismos actuales, constituyen el registro morfológico o anatómico. Al comparar las anatomías de especies modernas y extintas, los paleontólogos pueden inferir los linajes de esas especies. Sin embargo, este enfoque es más exitoso para organismos que tenían partes del cuerpo duras, como conchas, huesos o dientes. Además, como los procariotas, como las bacterias y las arqueas, comparten un conjunto limitado de morfologías comunes, sus fósiles no proporcionan información sobre su ascendencia.
Los procariotas habitaron la Tierra desde hace aproximadamente 3 a 4 mil millones de años. No se produjeron cambios obvios en la morfología o la organización celular en estos organismos durante los siguientes miles de millones de años. Las células eucariotas surgieron hace entre 1.6 y 2.7 mil millones de años. El siguiente cambio importante en la estructura celular se produjo cuando las bacterias fueron engullidas por células eucariotas, en una asociación cooperativa llamada endosimbiosis . Las bacterias engullidas y la célula huésped luego experimentaron coevolución, y las bacterias evolucionaron hacia mitocondrias o hidrogenosomas . Otro engullimiento de organismos parecidos a las cianobacterias condujo a la formación de cloroplastos en algas y plantas.


La historia de la vida fue la de los procariotas y eucariotas unicelulares hasta hace unos 610 millones de años cuando los organismos multicelulares comenzaron a aparecer en los océanos en el período ediacariano . La evolución de la multicelularidad ocurrió en múltiples eventos independientes, en organismos tan diversos como esponjas , algas pardas , cianobacterias , mohos limosos y mixobacterias . En 2016, los científicos informaron que, hace unos 800 millones de años, un cambio genético menor en una sola molécula llamada GK-PID pudo haber permitido que los organismos pasaran de un organismo de una sola célula a una de muchas células.
Poco después de la aparición de estos primeros organismos multicelulares, apareció una cantidad notable de diversidad biológica en un lapso de aproximadamente 10 millones de años, en un evento llamado explosión cámbrica . Aquí, la mayoría de los tipos de animales modernos aparecieron en el registro fósil, así como linajes únicos que posteriormente se extinguieron. Se han propuesto varios factores desencadenantes de la explosión del Cámbrico, incluida la acumulación de oxígeno en la atmósfera a partir de la fotosíntesis.
Fondo

Las palabras procariota y eucariota provienen del griego, donde pro significa "antes", eu significa "bien" o "verdadero", y karyon significa "nuez", "núcleo" o "núcleo". Entonces, etimológicamente, procariota significa "antes del núcleo" y eucariota significa "núcleo verdadero".
La división de las formas de vida entre procariotas y eucariotas fue establecida firmemente por los microbiólogos Roger Stanier y CB van Niel en su artículo de 1962, El concepto de bacteria . Una de las razones de esta clasificación fue que lo que entonces a menudo se llamaba algas verde-azuladas (ahora llamadas cianobacterias ) dejaría de clasificarse como plantas y se agruparía con bacterias.
En 1990 Carl Woese et al. introdujo el sistema de tres dominios . Los procariotas se dividieron en dos dominios , las arqueas y las bacterias, mientras que los eucariotas se convirtieron en un dominio por derecho propio. La diferencia clave con las clasificaciones anteriores es la separación de arqueas de bacterias.



La evidencia más temprana de vida en la tierra proviene de firmas de carbono biogénico y fósiles de estromatolitos descubiertos en rocas de 3.700 millones de años. En 2015, se encontraron posibles "restos de vida biótica " en rocas de 4.100 millones de años. En 2017, se reportó evidencia putativa de posiblemente las formas de vida más antiguas en la Tierra en forma de microorganismos fosilizados descubiertos en precipitados de respiraderos hidrotermales que pueden haber vivido tan pronto como hace 4.280 millones de años, no mucho después de que los océanos se formaran hace 4.400 millones de años, y no mucho después de la formación de la Tierra hace 4.540 millones de años.
Las esteras microbianas de bacterias y arqueas coexistentes eran la forma de vida dominante en el Eón Arcaico temprano y se cree que muchos de los pasos principales en la evolución temprana tuvieron lugar en este entorno. La evolución de la fotosíntesis alrededor de 3.5 Ga resultó en una acumulación de su producto de desecho de oxígeno en la atmósfera, lo que llevó al gran evento de oxigenación que comenzó alrededor de 2.4 Ga.
La evidencia más temprana de eucariotas data de 1.85 Ga, y aunque pueden haber estado presentes antes, su diversificación se aceleró cuando comenzaron a usar oxígeno en su metabolismo . Más tarde, alrededor de 1.7 Ga, comenzaron a aparecer organismos multicelulares , con células diferenciadas que realizaban funciones especializadas.

Una corriente de microorganismos transportados por el aire, incluidos los procariotas, rodea el planeta por encima de los sistemas meteorológicos pero por debajo de las rutas aéreas comerciales. Algunos microorganismos peripatéticos son arrastrados por las tormentas de polvo terrestres, pero la mayoría se origina a partir de microorganismos marinos en el rocío del mar . En 2018, los científicos informaron que cientos de millones de virus y decenas de millones de bacterias se depositan diariamente en cada metro cuadrado del planeta.
La vida microscópica submarina es diversa y aún no se comprende bien, como el papel de los virus en los ecosistemas marinos. La mayoría de los virus marinos son bacteriófagos , que son inofensivos para las plantas y los animales, pero son esenciales para la regulación de los ecosistemas de agua dulce y salada. Infectan y destruyen bacterias y arqueas en comunidades microbianas acuáticas y son el mecanismo más importante de reciclaje de carbono en el medio marino. Las moléculas orgánicas liberadas de las células bacterianas muertas estimulan el crecimiento de algas y bacterias frescas. La actividad viral también puede contribuir a la bomba biológica , el proceso por el cual el carbono se secuestra en las profundidades del océano.
Bacterias marinas




Las bacterias constituyen un gran dominio de microorganismos procarióticos . Por lo general, de unos pocos micrómetros de longitud, las bacterias tienen varias formas, que van desde esferas hasta varillas y espirales. Las bacterias estuvieron entre las primeras formas de vida que aparecieron en la Tierra y están presentes en la mayoría de sus hábitats . Las bacterias habitan en el suelo, el agua, las aguas termales ácidas , los desechos radiactivos y las partes profundas de la corteza terrestre . Las bacterias también viven en relaciones simbióticas y parasitarias con plantas y animales.
Una vez consideradas plantas que constituyen la clase Schizomycetes , las bacterias ahora se clasifican como procariotas . A diferencia de las células de animales y otros eucariotas , las células bacterianas no contienen un núcleo y rara vez albergan orgánulos unidos a la membrana . Aunque el término bacteria incluía tradicionalmente a todos los procariotas, la clasificación científica cambió después del descubrimiento en la década de 1990 de que los procariotas consisten en dos grupos muy diferentes de organismos que evolucionaron a partir de un ancestro común antiguo. Estos dominios evolutivos se denominan bacterias y arqueas .
Los antepasados de las bacterias modernas fueron microorganismos unicelulares que fueron las primeras formas de vida que aparecieron en la Tierra, hace unos 4 mil millones de años. Durante unos 3.000 millones de años, la mayoría de los organismos fueron microscópicos y las bacterias y las arqueas fueron las formas de vida dominantes. Aunque existen fósiles bacterianos , como los estromatolitos , su falta de morfología distintiva impide que se utilicen para examinar la historia de la evolución bacteriana o para fechar el momento de origen de una especie bacteriana en particular. Sin embargo, las secuencias de genes se pueden utilizar para reconstruir la filogenia bacteriana , y estos estudios indican que las bacterias divergieron primero del linaje archaeal / eucariota. Las bacterias también participaron en la segunda gran divergencia evolutiva, la de las arqueas y los eucariotas. Aquí, los eucariotas resultaron de la entrada de bacterias antiguas en asociaciones endosimbióticas con los antepasados de las células eucariotas, que posiblemente estaban relacionadas con las arqueas . Esto implicó la absorción por las células proto-eucariotas de simbiontes alfaproteobacterianos para formar mitocondrias o hidrogenosomas , que todavía se encuentran en todos los Eukarya conocidos. Más tarde, algunos eucariotas que ya contenían mitocondrias también engullieron organismos parecidos a las cianobacterias. Esto condujo a la formación de cloroplastos en algas y plantas. También hay algunas algas que se originaron incluso a partir de eventos endosimbióticos posteriores. Aquí, los eucariotas envolvieron un alga eucariota que se convirtió en un plastidio de "segunda generación". Esto se conoce como endosimbiosis secundaria .
Las bacterias crecen hasta un tamaño fijo y luego se reproducen mediante fisión binaria , una forma de reproducción asexual . En condiciones óptimas, las bacterias pueden crecer y dividirse extremadamente rápido, y las poblaciones bacterianas pueden duplicarse cada 9,8 minutos.
Pelagibacter ubique y sus parientes pueden ser los microorganismos más abundantes en el océano, y se ha afirmado que posiblemente sean las bacterias más abundantes del mundo. Constituyen aproximadamente el 25% de todas lascélulas de plancton microbiano, y en el verano pueden representar aproximadamente la mitad de las células presentes en el agua superficial del océano templado. Seestima que laabundancia total de P. ubique y sus parientes es de aproximadamente 2 × 10 28 microbios. Sin embargo, se informó en Nature en febrero de 2013 que el bacteriófago HTVC010P , que ataca a P. ubique , ha sido descubierto y es probablemente el organismo más común en el planeta.
Roseobacter es también uno de los microorganismos más abundantes y versátiles del océano. Están diversificados en diferentes tipos de hábitats marinos, desde la costa hasta los océanos abiertos y desde el hielo marino hasta el fondo del mar, y constituyen aproximadamente el 25% de las bacterias marinas costeras. Los miembros delgénero Roseobacter desempeñan un papel importante en los ciclos biogeoquímicos marinos y el cambio climático, procesando una parte significativa del carbono total en el medio marino. Forman relaciones simbióticas que les permiten degradar compuestos aromáticos y absorber trazas de metales. Se utilizan ampliamente en acuicultura y detección de quórum. Durante las floraciones de algas, el 20-30% de la comunidad procariota son Roseobacter.
La bacteria más grande conocida, la Thiomargarita namibiensis marina , puede ser visible a simple vista y en ocasiones alcanza 0,75 mm (750 μm).


Cianobacterias

.jpg)

Las cianobacterias fueron los primeros organismos en desarrollar la capacidad de convertir la luz solar en energía química. Forman un filo (división) de bacterias que van de unicelulares a filamentosas e incluyen especies coloniales . Se encuentran en casi todas partes de la tierra: en suelos húmedos, tanto en agua dulce como en ambientes marinos, e incluso en rocas antárticas. En particular, algunas especies se presentan como células a la deriva que flotan en el océano y, como tales, se encuentran entre las primeras del fitoplancton .
Los primeros productores primarios que utilizaron la fotosíntesis fueron las cianobacterias oceánicas hace unos 2.300 millones de años. La liberación de oxígeno molecular por las cianobacterias como subproducto de la fotosíntesis indujo cambios globales en el medio ambiente de la Tierra. Debido a que el oxígeno era tóxico para la mayoría de la vida en la Tierra en ese momento, esto llevó a la casi extinción de los organismos intolerantes al oxígeno , un cambio dramático que redirigió la evolución de las principales especies animales y vegetales.

interactúan con bacterias para adquirir hierro del polvo

Floración de las cianobacterias filamentosas Trichodesmium

Las floraciones de cianobacterias pueden contener cianotoxinas letales



Synechococcus , una cianobacteria marina muy extendida

Carboxisomas que aparecen como estructuras oscuras poliédricas dentro de una especie de Synechococcus



La diminuta cianobacteria marina Prochlorococcus (0,6 µm ) , descubierta en 1986, forma hoy una parte importante de la base de la cadena alimentaria del océano y representa gran parte de la fotosíntesis del océano abierto y se estima que el 20% del oxígeno en la atmósfera de la Tierra. . Es posiblemente el género más abundante en la Tierra: un solo mililitro de agua de mar superficial puede contener 100.000 células o más.
Originalmente, los biólogos clasificaron a las cianobacterias como algas y las denominaron "algas verdiazules". La opinión más reciente es que las cianobacterias son bacterias y, por lo tanto, ni siquiera están en el mismo reino que las algas. La mayoría de las autoridades excluyen a todos los procariotas y, por lo tanto, a las cianobacterias de la definición de algas.
| Video externo | |
|---|---|
|
|
Otras bacterias
Otras bacterias marinas, además de las cianobacterias, son ubicuas o pueden desempeñar un papel importante en el océano. Estos incluyen el copiotrofo oportunista , Alteromonas macleodii .
Arqueas marinas

Las arqueas (del griego antiguo ) constituyen un dominio y reino de microorganismos unicelulares . Estos microbios son procariotas , lo que significa que no tienen núcleo celular ni ningún otro orgánulo unido a la membrana en sus células.
Las arqueas se clasificaron inicialmente como bacterias , pero esta clasificación está desactualizada. Las células de Archaeal tienen propiedades únicas que las separan de los otros dos dominios de la vida, Bacteria y Eukaryota . Las arqueas se dividen además en múltiples filos reconocidos . La clasificación es difícil porque la mayoría no se han aislado en el laboratorio y solo se han detectado mediante el análisis de sus ácidos nucleicos en muestras de su entorno.
Las bacterias y las arqueas son generalmente similares en tamaño y forma, aunque algunas arqueas tienen formas muy extrañas, como las células planas y cuadradas de Haloquadratum walsbyi . A pesar de esta similitud morfológica con las bacterias, las arqueas poseen genes y varias vías metabólicas que están más estrechamente relacionadas con las de los eucariotas, en particular las enzimas involucradas en la transcripción y traducción . Otros aspectos de la bioquímica de las arqueas son únicos, como su dependencia de los éter lípidos en sus membranas celulares , como los arqueoles . Las arqueas utilizan más fuentes de energía que los eucariotas: estos van desde compuestos orgánicos , como azúcares, hasta amoníaco , iones metálicos o incluso gas hidrógeno . Las arqueas tolerantes a la sal (las Haloarchaea ) utilizan la luz solar como fuente de energía y otras especies de arqueas fijan el carbono ; sin embargo, a diferencia de las plantas y las cianobacterias , ninguna especie conocida de arqueas hace ambas cosas. Las arqueas se reproducen asexualmente por fisión binaria , fragmentación o gemación ; a diferencia de las bacterias y eucariotas, ninguna especie conocida forma esporas .
Las arqueas son particularmente numerosas en los océanos, y las arqueas del plancton pueden ser uno de los grupos de organismos más abundantes del planeta. Las arqueas son una parte importante de la vida de la Tierra y pueden desempeñar un papel tanto en el ciclo del carbono como en el ciclo del nitrógeno . Las crenarchaeota (eocitos) son un filo de arqueas que se cree que son muy abundantes en ambientes marinos y uno de los principales contribuyentes a la fijación de carbono.

Los eocitos pueden ser las arqueas marinas más abundantes

Las halobacterias , que se encuentran en el agua casi saturada de sal, ahora se reconocen como arqueas.

Células planas y cuadradas de las arqueas Haloquadratum walsbyi

Methanosarcina barkeri , una arquea marina que produce metano

Los termófilos , como Pyrolobus fumarii , sobreviven más de 100 ° C






Nanoarchaeum equitans es una especie de arqueas marinas descubierta en 2002 en un respiradero hidrotermal . Es un termófilo que crece a temperaturas de unos 80 grados centígrados. Nanoarchaeum parece ser un simbionte obligado en el archaeon Ignicoccus . Debe permanecer en contacto con el organismo huésped para sobrevivir, ya que Nanoarchaeum equitans no puede sintetizar lípidos, pero los obtiene de su huésped. Sus células tienen solo 400 nm de diámetro, lo que lo convierte en uno de los organismos celulares más pequeños conocidos y el arqueón más pequeño conocido.
Las arqueas marinas se han clasificado de la siguiente manera:
- Grupo marino I (MG-I o MGI): Thaumarchaeota marina con subgrupos Ia (también conocido como Ia) hasta Id
- Grupo marino II (MG-II): Euryarchaeota marina , orden Poseidoniales con subgrupos IIa hasta IId (IIa parecido a Poseidoniaceae , IIb parecido a Thalassarchaceae )
Los virus que parasitan MGII se clasifican como magrovirus - Marine Group III (MG-III): también marina Euryarchaeota, Marine Benthic Group D
- Marine Group IV (MG-IV): también marina Euryarchaeota
Modo trófico
El metabolismo de los procariotas se clasifica en grupos nutricionales sobre la base de tres criterios principales: la fuente de energía , los donantes de electrones utilizados y la fuente de carbono utilizada para el crecimiento.
| Tipo nutricional | Fuente de energía | Fuente de carbono | Ejemplos de |
|---|---|---|---|
| Fotótrofos | Luz del sol | Compuestos orgánicos (fotoheterótrofos) o fijación de carbono (fotoautótrofos) | Cianobacterias , bacterias verdes de azufre , cloroflexi o bacterias púrpuras |
| Litótrofos | Compuestos inorgánicos | Compuestos orgánicos (litoheterótrofos) o fijación de carbono (litoautótrofos) | Termodesulfobacterias , Hydrogenophilaceae o Nitrospirae |
| Organotrofos | Compuestos orgánicos | Compuestos orgánicos (quimioheterótrofos) o fijación de carbono (quimioautótrofos) | Bacillus , Clostridium o Enterobacteriaceae |
Los procariotas marinos se han diversificado enormemente a lo largo de su larga existencia. El metabolismo de los procariotas es mucho más variado que el de los eucariotas, lo que da lugar a muchos tipos de procariotas muy distintos. Por ejemplo, además de utilizar la fotosíntesis o los compuestos orgánicos para obtener energía, como hacen los eucariotas, los procariotas marinos pueden obtener energía de compuestos inorgánicos como el sulfuro de hidrógeno . Esto permite que los procariotas marinos prosperen como extremófilos en entornos hostiles tan fríos como la superficie del hielo de la Antártida, estudiada en criobiología , tan calientes como los respiraderos hidrotermales submarinos , o en condiciones de alta salinidad como los ( halófilos ). Algunos procariotas marinos viven simbióticamente en o sobre los cuerpos de otros organismos marinos.

- La fototrofia es un marcador particularmente significativo que siempre debe jugar un papel principal en la clasificación bacteriana.
- Las bacterias fototróficas aeroxigenicas (AAPB) son plancton marino ampliamente distribuido que pueden constituir más del 10% de la comunidad microbiana del océano abierto. Los AAPB marinos se clasifican en dos géneros marinos ( Erythrobacter y Roseobacter ). Pueden ser particularmente abundantes en condiciones oligotróficas donde se encontró que eran el 24% de la comunidad. Estos son organismos heterótrofos que utilizan la luz para producir energía, pero no pueden utilizar el dióxido de carbono como su principal fuente de carbono. La mayoría son estrictamente aeróbicos , lo que significa que necesitan oxígeno para crecer. Los datos actuales sugieren que las bacterias marinas tienen tiempos de generación de varios días, mientras que existe nueva evidencia que muestra que AAPB tiene un tiempo de generación mucho más corto. Las aguas costeras / de la plataforma a menudo tienen mayores cantidades de AAPB, algunas de hasta 13,51% AAPB%. El fitoplancton también afecta al AAPB%, pero se han realizado pocas investigaciones en esta área. También pueden ser abundantes en diversas condiciones oligotróficas , incluido el régimen más oligotrófico del océano mundial. Están distribuidos globalmente en la zona eufótica y representan un componente hasta ahora no reconocido de la comunidad microbiana marina que parece ser crítico para el ciclo del carbono orgánico e inorgánico en el océano.
- Bacterias moradas :
- Zetaproteobacteria : son quimiolitoautótrofos neutrófilos oxidantes del hierro , distribuidos mundialmente en estuarios y hábitats marinos.
- Las bacterias oxidantes de hidrógeno son autótrofas facultativas que se pueden dividir en aerobios y anaerobios. Los primeros usan hidrógeno como donante de electrones y oxígeno como aceptor, mientras que los segundos usan sulfato o dióxido de nitrógeno como aceptores de electrones .
Motilidad

La motilidad es la capacidad de un organismo para moverse de forma independiente, utilizando energía metabólica.
Motilidad flagelar


Los procariotas, tanto bacterias como arqueas, utilizan principalmente flagelos para la locomoción.
- Los flagelos bacterianos son filamentos helicoidales, cada uno con un motor rotatorio en su base que puede girar en sentido horario o antihorario. Proporcionan dos de varios tipos de motilidad bacteriana.
- Los flagelos arqueales se denominan archaella y funcionan de manera muy similar a los flagelos bacterianos. Estructuralmente, el archaellum es superficialmente similar a un flagelo bacteriano, pero difiere en muchos detalles y se considera no homólogo .
El modelo de motor rotatorio utilizado por las bacterias utiliza los protones de un gradiente electroquímico para mover sus flagelos. El torque en los flagelos de bacterias es creado por partículas que conducen protones alrededor de la base del flagelo. La dirección de rotación de los flagelos en las bacterias proviene de la ocupación de los canales de protones a lo largo del perímetro del motor flagelar.
Algunas células eucariotas también usan flagelos, y se pueden encontrar en algunos protistas y plantas, así como en células animales. Los flagelos eucariotas son proyecciones celulares complejas que se mueven hacia adelante y hacia atrás, en lugar de realizar un movimiento circular. Los flagelos procarióticos utilizan un motor rotatorio y los flagelos eucarióticos utilizan un complejo sistema de filamentos deslizantes. Los flagelos eucariotas están impulsados por ATP , mientras que los flagelos procariotas pueden ser impulsados por ATP (arqueas) o impulsados por protones (bacterias).
| Video externo | |
|---|---|
|
|
Motilidad espasmódica
La motilidad espasmódica es una forma de motilidad bacteriana rastrera que se utiliza para moverse sobre superficies. La contracción está mediada por la actividad de filamentos similares a pelos llamados pili tipo IV que se extienden desde el exterior de la célula, se unen a los sustratos sólidos circundantes y se retraen, tirando de la célula hacia adelante de una manera similar a la acción de un gancho de agarre . El nombre de motilidad espasmódica se deriva de los movimientos espasmódicos e irregulares característicos de las células individuales cuando se observan bajo el microscopio.
Motilidad deslizante
La motilidad de deslizamiento es un tipo de translocación que es independiente de estructuras propulsoras como flagelos o pili . El deslizamiento permite que los microorganismos viajen a lo largo de la superficie de películas poco acuosas. Los mecanismos de esta motilidad se conocen solo parcialmente. La velocidad de deslizamiento varía entre organismos, y la inversión de dirección aparentemente está regulada por algún tipo de reloj interno. Por ejemplo, los apicomplejos pueden viajar a velocidades rápidas entre 1 y 10 μm / s. Por el contrario, las bacterias Myxococcus xanthus se deslizan a una velocidad de 5 μm / min.
Motilidad enjambre
La motilidad de enjambre es una translocación rápida (2 a 10 μm / s) y coordinada de una población bacteriana a través de superficies sólidas o semisólidas, y es un ejemplo de multicelularidad bacteriana y comportamiento de enjambre . La motilidad del enjambre fue descrita por primera vez en 1972 por Jorgen Henrichsen.
Inmóvil
Las especies inmóviles carecen de la capacidad y las estructuras que les permitirían impulsarse, por su propio poder, a través de su entorno. Cuando se cultivan bacterias inmóviles en un tubo de apuñalamiento, solo crecen a lo largo de la línea de apuñalamiento. Si las bacterias son móviles, la línea aparecerá difusa y se extenderá hacia el medio.
Taxis: movimiento dirigido
Magnetotaxis


Las bacterias magnetotácticas se orientan a lo largo de las líneas del campo magnético del campo magnético de la Tierra . Se cree que esta alineación ayuda a estos organismos a alcanzar regiones de concentración óptima de oxígeno. Para realizar esta tarea, estas bacterias tienen orgánulos biomineralizados llamados magnetosomas que contienen cristales magnéticos . El fenómeno biológico de los microorganismos que tienden a moverse en respuesta a las características magnéticas del entorno se conoce como magnetotaxis . Sin embargo, este término es engañoso en el sentido de que cualquier otra aplicación del término taxis implica un mecanismo de estímulo-respuesta. En contraste con la magnetorrecepción de los animales, las bacterias contienen imanes fijos que fuerzan a las bacterias a alinearse, incluso las células muertas se alinean, como la aguja de una brújula.
Los ambientes marinos se caracterizan generalmente por bajas concentraciones de nutrientes mantenidos en movimiento constante o intermitente por corrientes y turbulencias. Las bacterias marinas han desarrollado estrategias, como la natación y el uso de sistemas de detección y respuesta direccionales, para migrar hacia lugares favorables en los gradientes de nutrientes. Las bacterias magnetotácticas utilizan el campo magnético de la Tierra para facilitar el nado descendente hacia la interfaz óxico-anóxica, que es el lugar más favorable para su persistencia y proliferación, en sedimentos o columnas de agua estratificados químicamente.

Dependiendo de su latitud y de si las bacterias están al norte o al sur del ecuador, el campo magnético de la Tierra tiene una de las dos posibles polaridades y una dirección que apunta con diferentes ángulos hacia las profundidades del océano y lejos de la superficie generalmente más rica en oxígeno. . La aerotaxis es la respuesta por la cual las bacterias migran a una concentración de oxígeno óptima en un gradiente de oxígeno. Varios experimentos han demostrado claramente que la magnetotaxis y la aerotaxis funcionan en conjunto en las bacterias magnetotácticas. Se ha demostrado que, en las gotas de agua, las bacterias magnetotácticas que nadan en un solo sentido pueden invertir su dirección de natación y nadar hacia atrás en condiciones reductoras ( concentración de oxígeno inferior a la óptima ), a diferencia de las condiciones óxicas (concentración de oxígeno superior a la óptima).
Independientemente de su morfología, todas las bacterias magnetotácticas estudiadas hasta ahora son móviles por medio de flagelos. Las bacterias magnetotácticas marinas, en particular, tienden a poseer un aparato flagelar elaborado que puede involucrar hasta decenas de miles de flagelos. Sin embargo, a pesar de una extensa investigación en los últimos años, aún no se ha establecido si las bacterias magnetotácticas dirigen sus motores flagelares en respuesta a su alineación en los campos magnéticos. Se ha propuesto la simbiosis con bacterias magnetotácticas como explicación de la magnetorrecepción en algunos protistas marinos . Se está investigando si una relación similar puede ser la base de la magnetorrecepción también en los vertebrados . Los magnetofósiles inequívocos más antiguos provienen de los lechos de cretáceo del sur de Inglaterra, aunque informes menos seguros de magnetofósiles se extienden a Gunflint Chert de 1.900 millones de años .
Vacuolas de gas
| Parte de una serie sobre |
| Plancton |
|---|
 |
Algunos procariotas marinos poseen vacuolas de gas . Las vacuolas de gas son nanocompartimentos libremente permeables al gas que permiten que las bacterias marinas y las arqueas controlen su flotabilidad . Toman la forma de vesículas unidas a membranas en forma de huso y se encuentran en algunos procariotas de plancton , incluidas algunas cianobacterias . Se necesita una flotabilidad positiva para mantener las células en los tramos superiores de la columna de agua, de modo que puedan continuar realizando la fotosíntesis . Las vacuolas de gas están formadas por una capa de proteína que tiene una superficie interna altamente hidrófoba , lo que la hace impermeable al agua (y evita que el vapor de agua se condense en el interior) pero permeable a la mayoría de los gases . Debido a que la vesícula de gas es un cilindro hueco, es probable que colapse cuando aumenta la presión circundante . La selección natural ha ajustado la estructura de la vesícula de gas para maximizar su resistencia al pandeo , incluida una proteína de refuerzo externa, GvpC, más bien como el hilo verde en una manguera trenzada. Existe una relación simple entre el diámetro de la vesícula de gas y la presión a la que colapsará: cuanto más ancha es la vesícula de gas, más débil se vuelve. Sin embargo, las vesículas de gas más anchas son más eficientes y proporcionan más flotabilidad por unidad de proteína que las vesículas de gas estrechas. Diferentes especies producen vesículas de gas de diferente diámetro, lo que les permite colonizar diferentes profundidades de la columna de agua (especies altamente competitivas de crecimiento rápido con vesículas de gas anchas en la mayoría de las capas superiores; especies de crecimiento lento, adaptadas a la oscuridad, con vesículas de gas estrechas y fuertes en las capas más profundas).
La célula alcanza su altura en la columna de agua sintetizando vesículas de gas. A medida que la célula se eleva, puede aumentar su carga de carbohidratos a través del aumento de la fotosíntesis. Demasiado alto y la célula sufrirá fotoblanqueo y posible muerte, sin embargo, el carbohidrato producido durante la fotosíntesis aumenta la densidad de la célula, provocando su hundimiento. El ciclo diario de acumulación de carbohidratos a partir de la fotosíntesis y el catabolismo de carbohidratos durante las horas oscuras es suficiente para ajustar la posición de la célula en la columna de agua, llevarla hacia la superficie cuando sus niveles de carbohidratos son bajos y necesita realizar la fotosíntesis, y permitir que se hunda lejos de la dañina radiación ultravioleta cuando los niveles de carbohidratos de la célula se hayan recuperado. Un exceso extremo de carbohidratos provoca un cambio significativo en la presión interna de la célula, lo que hace que las vesículas de gas se doblen y colapsen y la célula se hunda.
Las vacuolas grandes se encuentran en tres géneros de bacterias de azufre filamentosas , Thioploca , Beggiatoa y Thiomargarita . El citosol es extremadamente reducido en estos géneros y la vacuola puede ocupar entre el 40 y el 98% de la célula. La vacuola contiene altas concentraciones de iones nitrato y, por lo tanto, se cree que es un orgánulo de almacenamiento .
Bioluminiscencia


Bacterias bioluminiscentes son productoras de luz bacterias que son predominantemente presente en agua de mar, sedimentos marinos, la superficie de la descomposición de pescado y en el intestino de los animales marinos. Aunque no es tan común, la bioluminiscencia bacteriana también se encuentra en bacterias terrestres y de agua dulce. Estas bacterias pueden ser de vida libre (como Vibrio harveyi ) o en simbiosis con animales como el calamar Bobtail hawaiano ( Aliivibrio fischeri ) o nematodos terrestres ( Photorhabdus luminescens ). Los organismos huéspedes proporcionan a estas bacterias un hogar seguro y una nutrición suficiente. A cambio, los anfitriones utilizan la luz producida por las bacterias para camuflarse, atraer presas y / o parejas. Las bacterias bioluminiscentes han desarrollado relaciones simbióticas con otros organismos en los que ambos participantes se benefician casi por igual. Otra posible razón por la que las bacterias usan la reacción de luminiscencia es para la detección de quórum , una capacidad para regular la expresión génica en respuesta a la densidad de células bacterianas.
El calamar bobtail hawaiano vive en simbiosis con la bacteria bioluminiscente Aliivibrio fischeri, que habita un órgano de luz especial en el manto del calamar. Las bacterias son alimentadas con azúcar y aminoácidos por el calamar y, a cambio, ocultan la silueta del calamar cuando se ve desde abajo, iluminándola contrariamente al igualar la cantidad de luz que llega a la parte superior del manto. El calamar sirve como organismo modelo para la simbiosis animal-bacteriana y su relación con la bacteria ha sido ampliamente estudiada.
Vibrio harveyi es unabacteria bioluminiscentemóvil en forma de varilla (a través de flagelos polares) que crece de manera óptima entre 30 ° y 35 ° C. Se puede encontrar nadando libremente en aguas marinas tropicales, comensalmente en la microflora intestinal deanimales marinos y como patógeno primario y oportunista de varios animales marinos. Se cree que es la causa del efecto de los mares lechosos , donde se emite un resplandor azul uniforme del agua de mar durante la noche. Algunos resplandores pueden cubrir casi 6.000 millas cuadradas (16.000 km 2 ).
Rodopsina microbiana

(2) cambia su configuración de modo que un protón es expulsado de la célula
(3) el potencial químico hace que el protón regrese a la célula
(4) generando energía
(5) en el forma de trifosfato de adenosina .
El metabolismo fototrófico se basa en uno de los tres pigmentos de conversión de energía: clorofila , bacterioclorofila y retina . La retina es el cromóforo que se encuentra en las rodopsinas . Durante décadas se ha escrito sobre la importancia de la clorofila en la conversión de energía luminosa, pero la fototrofia basada en pigmentos retinianos apenas está comenzando a estudiarse.
.jpg)
En 2000, un equipo de microbiólogos dirigido por Edward DeLong hizo un descubrimiento crucial en la comprensión de los ciclos de energía y carbono marino. Descubrieron un gen en varias especies de bacterias responsables de la producción de la proteína rodopsina , algo nunca antes visto en las bacterias. Estas proteínas que se encuentran en las membranas celulares son capaces de convertir la energía lumínica en energía bioquímica debido a un cambio en la configuración de la molécula de rodopsina cuando la luz solar la golpea, provocando el bombeo de un protón de adentro hacia afuera y una afluencia posterior que genera la energía. Las rodopsinas similares a arqueas se han encontrado posteriormente entre diferentes taxones, protistas, así como en bacterias y arqueas, aunque son raras en organismos multicelulares complejos .
La investigación en 2019 muestra que estas "bacterias que arrebatan el sol" están más extendidas de lo que se pensaba anteriormente y podrían cambiar la forma en que los océanos se ven afectados por el calentamiento global. "Los hallazgos rompen con la interpretación tradicional de la ecología marina que se encuentra en los libros de texto, que establece que casi toda la luz solar en el océano es capturada por la clorofila de las algas. En cambio, las bacterias equipadas con rodopsina funcionan como autos híbridos, alimentados por materia orgánica cuando está disponible, como la mayoría de las bacterias lo son, y por la luz solar cuando los nutrientes son escasos ".
Existe una conjetura astrobiológica llamada hipótesis de la Tierra Púrpura que supone que las formas de vida originales en la Tierra se basaban en la retina en lugar de la clorofila, lo que habría hecho que la Tierra pareciera púrpura en lugar de verde.
Simbiosis
Algunos organismos marinos tienen simbiosis con bacterias o arqueas. Los gusanos de Pompeya viven a grandes profundidades gracias a los respiraderos hidrotermales a temperaturas de hasta 80 ° C. Tienen lo que parecen ser espaldas peludas, pero estos "pelos" son en realidad colonias de bacterias como la Nautilia profundicola , que se cree que proporcionan al gusano cierto grado de aislamiento. Las glándulas de la espalda del gusano segregan un moco del que se alimentan las bacterias, una forma de simbiosis .

Hesiocaeca methanicola vive a grandes profundidades en el hielo de metano y parece sobrevivir en simbiosis con bacterias que metabolizan el clatrato .

Olavius algarvensis depende de cinco especies diferentes de bacterias simbióticas para su nutrición.
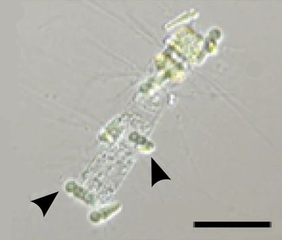
Cianobacterias epífitas de Calothrix (flechas) en simbiosis con una diatomea Chaetoceros . Barra de escala de 50 μm.





Las bacterias endosimbiontes son bacterias que viven dentro del cuerpo o en las células de otro organismo. Algunos tipos de cianobacterias son endosimbiontes y se ha descubierto que las cianobacterias poseen genes que les permiten someterse a la fijación de nitrógeno.
Los organismos suelen establecer una relación simbiótica debido a su limitada disponibilidad de recursos en su hábitat o debido a una limitación de su fuente de alimento. Las bacterias simbióticas y quimiosintéticas que se han descubierto asociadas con los mejillones ( Bathymodiolus ) ubicados cerca de los respiraderos hidrotermales tienen un gen que les permite utilizar hidrógeno como fuente de energía, en lugar de azufre o metano como fuente de energía para la producción de energía.
Olavius algarvensis es un gusano que vive en los sedimentos costeros del Mediterráneo y depende de bacterias simbióticas para su nutrición. Vive con cinco especies diferentes de bacterias ubicadas debajo de su cutícula: dos oxidantes de sulfuro , dos reductoras de sulfato y una espiroqueta . Las bacterias simbióticas también permiten que el gusano utilice hidrógeno y monóxido de carbono como fuentes de energía y metabolice compuestos orgánicos como malato y acetato .
Astrangia poculata , el coral estrella del norte, es un coral pedregoso templado, ampliamente documentado a lo largo de la costa este de los Estados Unidos. El coral puede vivir con y sin zooxantelas (simbiontes de algas), lo que lo convierte en un organismo modelo idealpara estudiar las interacciones de la comunidad microbiana asociadas con el estado simbiótico. Sin embargo, la capacidad de desarrollar cebadores y sondas para apuntar más específicamente a grupos microbianos clave se ha visto obstaculizada por la falta de secuencias de ARNr 16S de longitud completa, ya que las secuencias producidas por la plataforma Illumina son de longitud insuficiente (aproximadamente 250 pares de bases) para el diseño de cebadores y sondas. En 2019, Goldsmith et al. demostró que la secuenciación de Sanger era capaz de reproducir la diversidad biológicamente relevante detectada por una secuenciación más profunda de próxima generación , mientras que también producía secuencias más largas útiles para la comunidad de investigadores para el diseño de sondas y cebadores (ver diagrama a la derecha).
Funciones en las redes tróficas marinas

La mayor parte del volumen del océano mundial está a oscuras. Los procesos que ocurren dentro de la delgada capa superficial iluminada (la capa fótica desde la superficie hasta entre 50 y 170 metros) son de gran importancia para la biosfera global. Por ejemplo, la región visible del espectro solar (la llamada radiación fotosintéticamente disponible o PAR) que alcanza esta capa iluminada por el sol alimenta aproximadamente la mitad de la productividad primaria del planeta y es responsable de aproximadamente la mitad del oxígeno atmosférico necesario para la mayor parte de la vida. en la tierra.
El bacterioplancton heterotrófico son los principales consumidores de materia orgánica disuelta (DOM) en las redes tróficas marinas pelágicas , incluidas las capas superiores del océano iluminadas por el sol. Su sensibilidad a la radiación ultravioleta (UVR), junto con algunos mecanismos recientemente descubiertos que las bacterias han desarrollado para beneficiarse de la radiación fotosintéticamente disponible (PAR), sugieren que la luz solar natural juega un papel relevante, aunque difícil de predecir, en la modulación de las funciones biogeoquímicas bacterianas en los océanos.
Los hábitats de la superficie del océano se encuentran en la interfaz entre la atmósfera y el océano. El hábitat similar a una biopelícula en la superficie del océano alberga microorganismos que habitan en la superficie, comúnmente conocidos como neuston . Esta vasta interfaz aire-agua se encuentra en la intersección de los principales procesos de intercambio aire-agua que abarcan más del 70% de la superficie mundial. Las bacterias en la microcapa superficial del océano, llamadas bacterioneuston , son de interés debido a aplicaciones prácticas como el intercambio de gases de efecto invernadero entre el aire y el mar, la producción de aerosoles marinos climáticamente activos y la teledetección del océano. De interés específico es la producción y degradación de tensioactivos (materiales tensioactivos) a través de procesos bioquímicos microbianos. Las principales fuentes de tensioactivos en el océano abierto incluyen el fitoplancton, la escorrentía terrestre y la deposición de la atmósfera.

A diferencia de las floraciones de algas coloreadas, es posible que las bacterias asociadas a los surfactantes no sean visibles en las imágenes en color del océano. Tener la capacidad de detectar estas bacterias "invisibles" asociadas a los surfactantes usando un radar de apertura sintética tiene inmensos beneficios en todas las condiciones climáticas, independientemente de las nubes, la niebla o la luz del día. Esto es particularmente importante en vientos muy fuertes, porque estas son las condiciones en las que tienen lugar los intercambios de gas aire-mar y la producción de aerosoles marinos más intensos. Por lo tanto, además de las imágenes satelitales en color, las imágenes satelitales SAR pueden proporcionar información adicional sobre una imagen global de los procesos biofísicos en el límite entre el océano y la atmósfera, los intercambios de gases de efecto invernadero aire-mar y la producción de aerosoles marinos climáticamente activos.

El diagrama de la derecha muestra los vínculos entre la bomba biológica del océano y la red trófica pelágica y la capacidad de muestrear estos componentes de forma remota desde barcos, satélites y vehículos autónomos. Las aguas azul claro son la zona eufótica , mientras que las aguas azules más oscuras representan la zona crepuscular .
Funciones en el ciclo biogeoquímico
Las arqueas reciclan elementos como el carbono , el nitrógeno y el azufre a través de sus diversos hábitats. Las arqueas llevan a cabo muchos pasos en el ciclo del nitrógeno . Esto incluye tanto las reacciones que eliminan el nitrógeno de los ecosistemas (como la respiración basada en nitratos y la desnitrificación ) como los procesos que introducen nitrógeno (como la asimilación y la fijación de nitrógeno ).
Los investigadores descubrieron recientemente la participación de las arqueas en las reacciones de oxidación del amoníaco . Estas reacciones son particularmente importantes en los océanos. En el ciclo del azufre , las arqueas que crecen al oxidar compuestos de azufre liberan este elemento de las rocas, poniéndolo a disposición de otros organismos, pero las arqueas que hacen esto, como Sulfolobus , producen ácido sulfúrico como producto de desecho y el crecimiento de estos organismos. en minas abandonadas puede contribuir al drenaje ácido de la mina y otros daños ambientales. En el ciclo del carbono , las arqueas del metanógeno eliminan el hidrógeno y juegan un papel importante en la descomposición de la materia orgánica por parte de las poblaciones de microorganismos que actúan como descomponedores en ecosistemas anaeróbicos, como sedimentos y marismas.
Ver también
- Métodos de recuento de bacterioplancton
- Bacterias bioluminiscentes
- Bacterias oxidantes de hierro
- Pelagibacterales - organismos modelo en teoría de racionalización
- Racionalización de la teoría
Referencias
Bibliografía
- Dalrymple, G. Brent (2001). "La edad de la Tierra en el siglo XX: un problema (en su mayoría) resuelto". Publicaciones especiales, Sociedad Geológica de Londres . 190 (1): 205–221. Código Bibliográfico : 2001GSLSP.190..205D . doi : 10.1144 / GSL.SP.2001.190.01.14 . S2CID 130092094 .
- Cuervo, Peter Hamilton; Johnson, George Brooks (2002). Biología . Educación McGraw-Hill. ISBN 978-0-07-112261-0.
- Shors, Teri (2017). Entender los virus . Jones y Bartlett Publishers. ISBN 978-1284025927.