
Concha protista - Protist shell

| Parte de una serie relacionada con |
| Biomineralización |
|---|
 |
Muchos protistas tienen carcasas o pruebas protectoras , generalmente hechas de sílice (vidrio) o carbonato de calcio (tiza). Los protistas son en su mayoría unicelulares y microscópicos. Sus caparazones son a menudo formas mineralizadas duras que resisten la degradación y pueden sobrevivir a la muerte del protista como un microfósil . Aunque los protistas son muy pequeños, son ubicuos. Su número es tal que sus conchas juegan un papel muy importante en la formación de sedimentos oceánicos y en el ciclo global de elementos y nutrientes.
Visión general
La vida se originó como procariotas unicelulares (bacterias y arqueas) y luego evolucionó a eucariotas más complejos . Los eucariotas son las formas de vida más desarrolladas conocidas como plantas, animales, hongos y protistas. Los protistas son los eucariotas que no se pueden clasificar como plantas, hongos o animales. Ha habido poca investigación sobre protistas mixotróficos , pero estudios recientes en ambientes marinos encontraron que los protistas mixotróficos contribuyen con una parte significativa de la biomasa protista . Los protistas suelen ser unicelulares y microscópicos. El término protista se utilizó históricamente como un término de conveniencia para eucariotas que no pueden clasificarse estrictamente como plantas, animales u hongos. No forman parte de la cladística moderna, porque son parafiléticos (carecen de un ancestro común).
La mayoría de los protistas son demasiado pequeños para ser vistos a simple vista. Son organismos muy diversos actualmente organizados en 18 filos, pero no fáciles de clasificar. Los estudios han demostrado que existe una alta diversidad de protistas en los océanos, los respiraderos de aguas profundas y los sedimentos de los ríos, lo que sugiere que aún no se han descubierto un gran número de comunidades microbianas eucariotas. Dado que los protistas son eucariotas (y no procariotas), poseen dentro de su célula al menos un núcleo , así como orgánulos como las mitocondrias y los cuerpos de Golgi . Los protistas son asexuales pero pueden reproducirse rápidamente por mitosis o por fragmentación .
A diferencia de las células de bacterias y arqueas, las células de protistas y otros eucariotas están muy organizadas. Las plantas, los animales y los hongos suelen ser multicelulares y típicamente macroscópicos . La mayoría de los protistas son unicelulares y microscópicos. Pero hay excepciones, y algunos protistas marinos no son unicelulares ni microscópicos, como las algas .
A base de silicio

Aunque el silicio está fácilmente disponible en forma de silicatos , muy pocos organismos lo utilizan directamente. Las diatomeas , radiolarios y esponjas silíceas utilizan sílice biogénica como material estructural para sus esqueletos. En plantas más avanzadas, los fitolitos de sílice ( fitolitos de ópalo) son cuerpos microscópicos rígidos que se encuentran en la célula; algunas plantas, por ejemplo el arroz , necesitan silicio para crecer. Se ha demostrado que el silicio mejora la resistencia de la pared celular de las plantas y la integridad estructural en algunas plantas.
Diatomeas
Las diatomeas forman un filo (disputado) que contiene alrededor de 100.000 especies reconocidas de algas principalmente unicelulares. Las diatomeas generan alrededor del 20 por ciento del oxígeno producido en el planeta cada año, absorben más de 6,7 mil millones de toneladas métricas de silicio cada año de las aguas en las que viven y contribuyen con casi la mitad del material orgánico que se encuentra en los océanos.
Las diatomeas están encerradas en conchas protectoras de sílice (vidrio) llamadas frústulas . La estructura intrincada y bellamente diseñada de muchas de estas frústulas es tal que a menudo se las conoce como "joyas del mar". Cada frústula está hecha de dos partes entrelazadas cubiertas con pequeños orificios a través de los cuales la diatomea intercambia nutrientes y desechos. Las frústulas de las diatomeas muertas se desplazan hasta el fondo del océano, donde, durante millones de años, pueden acumularse hasta un kilómetro de profundidad .
Las diatomeas utilizan silicio en forma de sílice biogénica (BSIO 2 ), que es absorbido por la proteína de transporte de silicio para ser utilizado predominantemente en la construcción de estas estructuras protectoras de la pared celular. El silicio ingresa al océano en forma disuelta, como ácido silícico o silicato . Dado que las diatomeas son uno de los principales usuarios de estas formas de silicio, contribuyen en gran medida a la concentración de silicio en todo el océano. El silicio forma un perfil similar a un nutriente en el océano debido a la productividad de las diatomeas en profundidades poco profundas, lo que significa que hay menos concentración de silicio en la parte superior del océano y más concentración de silicio en las profundidades del océano.
La productividad de las diatomeas en la parte superior del océano contribuye a la cantidad de silicio exportado a la parte inferior del océano. Cuando las células de diatomeas se lisan en la parte superior del océano, sus nutrientes, como el hierro, el zinc y el silicio, se llevan al océano inferior a través de un proceso llamado nieve marina . La nieve marina implica la transferencia descendente de materia orgánica particulada mediante la mezcla vertical de materia orgánica disuelta . La disponibilidad de silicio parece crucial para la productividad de las diatomeas, y siempre que el ácido silícico esté disponible para que lo utilicen las diatomeas, las diatomeas contribuyen con otras importantes concentraciones de nutrientes en las profundidades del océano.
En las zonas costeras, las diatomeas sirven como los principales organismos fitoplanctónicos y contribuyen en gran medida a la producción de sílice biogénica. En mar abierto, sin embargo, las diatomeas tienen un papel reducido en la producción anual mundial de sílice. Las diatomeas en los giros subtropicales del Atlántico Norte y el Pacífico Norte contribuyen solo con alrededor del 6% de la producción anual de sílice marina mundial, mientras que el Océano Austral produce alrededor de un tercio de la sílice biogénica marina mundial. Se dice que el Océano Austral tiene una "división biogeoquímica", ya que solo se transportan cantidades minúsculas de silicio fuera de esta región.



Las diatomeas son uno de los tipos más comunes de fitoplancton.

Sus conchas protectoras (frustles) están hechas de silicona.


Vienen en muchas formas y tamaños.
_Various_diatoms.jpg)



.jpg)
.jpg)

Frústula silicificada de una diatomea pennada con dos mitades superpuestas

Guinardia delicatula , una diatomea responsable de la proliferación de algas en el Mar del Norte y el Canal de la Mancha

Diatomea fósil

Hay más de 100.000 especies de diatomeas que representan el 50% de la producción primaria del océano.








Las frústulas de diatomeas se han acumulado durante más de 100 millones de años, dejando ricos depósitos de óxido de silicio nano y microestructurado en forma de tierra de diatomeas en todo el mundo. Las causas evolutivas de la generación de sílice nano y microestructurada por las algas fotosintéticas aún no están claras. Sin embargo, en 2018 se demostró que la reflexión de la luz ultravioleta por la sílice nanoestructurada protege el ADN en las células de las algas, y esto puede ser una causa evolutiva para la formación de las jaulas de vidrio.


| Video externo | |
|---|---|
|
| |
|
|
Radiolarios


Los radiolarios son protistas depredadores unicelulares encerrados en elaboradas conchas globulares generalmente hechas de sílice y perforadas con agujeros. Su nombre proviene del latín "radio". Atrapan a sus presas extendiendo partes de su cuerpo a través de los agujeros. Al igual que con las frústulas de sílice de las diatomeas, las conchas de radiolarios pueden hundirse en el fondo del océano cuando mueren los radiolarios y se conservan como parte del sedimento oceánico. Estos restos, como microfósiles , proporcionan información valiosa sobre las condiciones oceánicas pasadas.

Al igual que las diatomeas, los radiolarios tienen muchas formas.

También como las diatomeas, las conchas de radiolarios suelen estar hechas de silicato.

Sin embargo, los radiolarios acantários tienen conchas hechas de cristales de sulfato de estroncio.

.jpg)
.jpg)


Diagrama esquemático en corte de una concha radiolariana esférica

Cladococcus abietinus




.jpg)
| Video externo | |
|---|---|
|
| |
|
|
A base de calcio
Cocolitóforos
Los cocolitóforos son diminutos protistas fotosintéticos unicelulares con dos flagelos para la locomoción. La mayoría de ellos están protegidos por un caparazón llamado cocosfera . Las cocosferas están cubiertas con placas o escamas circulares ornamentadas llamadas cocolitos . Los cocolitos están hechos de carbonato de calcio. El término cocolitóforo deriva del griego para una piedra portadora de semillas , refiriéndose a su pequeño tamaño y las piedras de cocolito que llevan. En las condiciones adecuadas, florecen, como otros fitoplancton, y pueden convertir el océano en un blanco lechoso.


.png)


Existen beneficios para los protistas que llevan caparazones protectores. El diagrama de la izquierda a continuación muestra algunos de los beneficios que obtienen los cocolitóforos al ser portadores de cocolitos. En el diagrama, (A) representa la fotosíntesis acelerada, incluidos los mecanismos de concentración de carbono (CCM) y la captación de luz mejorada a través de la dispersión de fotones escasos para las especies que habitan en las profundidades. (B) representa la protección contra el fotodaño, incluida la protección del parasol contra la luz ultravioleta (UV) y la radiación fotosintética activa (PAR) y la disipación de energía en condiciones de mucha luz. (C) representa la protección de la armadura incluye protección contra infecciones virales / bacterianas y pastoreo por pastores selectivos y no selectivos.


También existen costos para los protistas que llevan carcasas protectoras. El diagrama de la derecha muestra algunos de los costos energéticos en los que incurren los cocolitóforos por transportar cocolitos. En el diagrama, los costos energéticos se informan en porcentaje del presupuesto fotosintético total. (A) representa que los procesos de transporte incluyen el transporte a la célula desde el agua de mar circundante de los sustratos de calcificación primaria Ca 2 + y HCO 3 - (flechas negras) y la eliminación del producto final H + de la célula (flecha gris). El transporte de Ca 2 + a través del citoplasma hasta la vesícula del cocolito (CV) es el costo dominante asociado con la calcificación. (B) representa los procesos metabólicos que incluyen la síntesis de polisacáridos asociados a cocolitos (CAP - rectángulos grises) por el complejo de Golgi (rectángulos blancos) que regulan la nucleación y geometría de los cristales de CaCO 3 . El cocolito completo (placa gris) es una estructura compleja de CAP y cristales de CaCO 3 intrincadamente dispuestos . (C) Los procesos mecánicos y estructurales explican la secreción de los cocolitos completos que se transportan desde su posición original adyacente al núcleo hasta la periferia celular, donde se transfieren a la superficie de la célula.
Foraminíferos


Al igual que los radiolarios, foraminíferos ( forams para abreviar) son depredadores protistas unicelulares, también protegidas con conchas que tienen agujeros en ellos. Su nombre proviene del latín para "portadores de agujeros". Sus conchas, a menudo llamadas pruebas , tienen cámaras (los foraminíferos agregan más cámaras a medida que crecen). Las conchas generalmente están hechas de calcita, pero a veces están hechas de partículas de sedimento aglutinadas o quitón , y (rara vez) de sílice. La mayoría de los foraminíferos son bentónicos, pero alrededor de 40 especies son plancticas. Se investigan ampliamente con registros fósiles bien establecidos que permiten a los científicos inferir mucho sobre ambientes y climas pasados.


| Video externo | |
|---|---|
|
| |
|
|


sección que muestra las cámaras de un foraminífero en espiral

Ectoplasma granular de transmisión de amoníaco tepida vivo para atrapar alimentos

Grupo de foraminíferos planctónicos

Fósiles nummulitid forams de varios tamaños, desde el Eoceno

Las pirámides egipcias se construyeron con piedra caliza que contenía nummulitas .





Otro
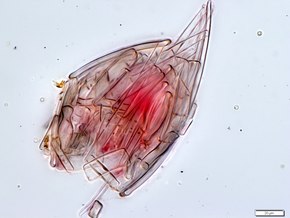
Loricifera (del latín lorica para corselet o armadura + ferre to bear) es un filo de animales marinos cicloneurales que habitan en sedimentos desde muy pequeños a microscópicos . Sus tamaños oscilan entre aproximadamente 100 μm y 1 mm. Se caracterizan por una carcasa exterior protectora llamada lorica y su hábitat está en los espacios entre la grava marina a la que se adhieren. La lorica con forma de armadura consiste en una capa externa protectora o una caja de plicae circundante .
El cuerpo celular de muchos coanoflagelados está rodeado por una matriz extracelular distintiva o periplasto . Estos recubrimientos celulares varían mucho en estructura y composición y los taxónomos los utilizan con fines de clasificación. Muchos coanoflagelados construyen "casas" complejas en forma de canasta, llamadas lorica , a partir de varias tiras de sílice cementadas entre sí. Se desconoce el significado funcional del periplasto, pero en los organismos sésiles se cree que ayuda a la unión al sustrato. En los organismos planctónicos, se especula que el periplasto aumenta la resistencia, contrarrestando así la fuerza generada por el flagelo y aumentando la eficiencia de la alimentación.
| Video externo | |
|---|---|
|
|



Microfósiles

Los microfósiles son fósiles que generalmente tienen entre 0,001 mm y 1 mm de tamaño, cuyo estudio requiere el uso de microscopía óptica o electrónica . Los fósiles que se pueden estudiar a simple vista o con un aumento de baja potencia, como una lupa, se denominan macrofósiles .
Por ejemplo, algunos organismos coloniales, como los Bryozoa (especialmente los Cheilostomata ) tienen colonias relativamente grandes , pero se clasifican por finos detalles esqueléticos de los pequeños individuos de la colonia.
En otro ejemplo, muchos géneros fósiles de Foraminifera , que son protistas, se conocen a partir de conchas (llamadas "pruebas") que eran tan grandes como monedas, como el género Nummulites .
Los microfósiles son una característica común del registro geológico , desde el Precámbrico hasta el Holoceno . Son más comunes en depósitos de ambientes marinos , pero también ocurren en agua salobre, agua dulce y depósitos sedimentarios terrestres . Si bien cada reino de la vida está representado en el registro de microfósiles, las formas más abundantes son los esqueletos protistas o quistes de Chrysophyta , Pyrrhophyta , Sarcodina , acritarchs y quitinozoans , junto con el polen y las esporas de las plantas vasculares .
En 2017, se anunció que se habían descubierto microorganismos fosilizados , o microfósiles, en los precipitados de respiraderos hidrotermales en el cinturón de Nuvvuagittuq de Quebec, Canadá, que puede tener hasta 4.280 millones de años, el registro más antiguo de vida en la Tierra , lo que sugiere "una casi emergencia instantánea de vida "(en un sentido de escala de tiempo geológico), después de la formación de los océanos hace 4.410 millones de años , y no mucho después de la formación de la Tierra hace 4.540 millones de años. No obstante, la vida puede haber comenzado incluso antes, hace casi 4.500 millones de años, como afirman algunos investigadores.
Ver también
Referencias
Otras referencias
- Xu, K., Hutchins, D. y Gao, K. (2018) "La disposición del cocolito sigue las matemáticas eulerianas en el cocolitóforo Emiliania huxleyi ". PeerJ , 6 : e4608. doi : 10.1126 / science.aaa7378 .
- Esqueletos de Protistán: una historia geológica de evolución y restricción