
Aminoácidos no proteinogénicos - Non-proteinogenic amino acids

En bioquímica , los aminoácidos no codificados o no proteinogénicos son distintos de los 22 aminoácidos proteinogénicos (21 en eucariotas) que están codificados naturalmente en el genoma de los organismos para el ensamblaje de proteínas. Sin embargo, más de 140 aminoácidos no proteinogénicos se encuentran naturalmente en las proteínas y miles más pueden ocurrir en la naturaleza o sintetizarse en el laboratorio. Muchos aminoácidos no proteinogénicos son importantes:
- intermedios en biosíntesis,
- en la formación postraduccional de proteínas,
- en un papel fisiológico (por ejemplo, componentes de las paredes celulares bacterianas , neurotransmisores y toxinas ),
- compuestos farmacológicos naturales o artificiales,
- presente en meteoritos o utilizado en experimentos prebióticos (por ejemplo, el experimento de Miller-Urey ).
Definición por negación

Técnicamente, cualquier compuesto orgánico con una amina (-NH 2 ) y un ácido carboxílico (-COOH) grupo funcional es un ácido amino. Los aminoácidos proteinogénicos son un pequeño subconjunto de este grupo que posee un átomo de carbono central (α- o 2-) que lleva un grupo amino, un grupo carboxilo, una cadena lateral y una conformación levo α-hidrógeno , con la excepción de la glicina , que es aquiral y prolina , cuyo grupo amina es una amina secundaria y, en consecuencia, se denomina frecuentemente iminoácido por razones tradicionales, aunque no como imino.
El código genético codifica 20 aminoácidos estándar para su incorporación en proteínas durante la traducción . Sin embargo, hay dos aminoácidos proteinogénicos adicionales: selenocisteína y pirrolisina . Estos aminoácidos no estándar no tienen un codón dedicado, pero se añaden en lugar de un codón de parada cuando está presente una secuencia específica, el codón UGA y el elemento SECIS para la selenocisteína, la secuencia aguas abajo UAG PYLIS para la pirrolisina. Todos los demás aminoácidos se denominan "no proteinogénicos".

Selenocisteína. Este aminoácido contiene un grupo selenol en su carbono β.

Pirrolisina. Este aminoácido se forma uniendo al grupo ε-amino de la lisina un anillo de pirrolina carboxilado.

Hay varios grupos de aminoácidos:
- 20 aminoácidos estándar
- 22 aminoácidos proteinogénicos
- más de 80 aminoácidos creados abióticamente en altas concentraciones
- alrededor de 900 son producidos por vías naturales
- más de 118 aminoácidos modificados se han colocado en proteínas
Estos grupos se superponen, pero no son idénticos. Los 22 aminoácidos proteinogénicos son biosintetizados por organismos y algunos, pero no todos, también son abióticos (que se encuentran en experimentos prebióticos y meteoritos). Algunos aminoácidos naturales, como la norleucina , están mal incorporados de forma traduccional en proteínas debido a la infidelidad del proceso de síntesis de proteínas. Muchos aminoácidos, como la ornitina , son intermediarios metabólicos producidos biosintéticamente, pero no incorporados por traducción en proteínas. La modificación postraduccional de residuos de aminoácidos en proteínas conduce a la formación de muchos aminoácidos proteínicos, pero no proteinogénicos. Otros aminoácidos se encuentran únicamente en mezclas abióticas (por ejemplo, α-metilnorvalina). Más de 30 aminoácidos no naturales se han insertado de forma traduccional en proteínas en sistemas diseñados, pero no son biosintéticos.
Nomenclatura
Además del sistema de numeración IUPAC para diferenciar los diversos carbonos en una molécula orgánica, asignando secuencialmente un número a cada carbono, incluidos los que forman un grupo carboxílico, los carbonos a lo largo de la cadena lateral de aminoácidos también se pueden marcar con letras griegas. , donde el carbono α es el carbono quiral central que posee un grupo carboxilo, una cadena lateral y, en los α-aminoácidos, un grupo amino; el carbono de los grupos carboxílicos no se cuenta. (En consecuencia, los nombres IUPAC de muchos α-aminoácidos no proteinogénicos comienzan con ácido 2-amino- y terminan en ácido -ico ).
Aminoácidos naturales, pero no L-α
La mayoría de los aminoácidos naturales son α-aminoácidos en la conformación L, pero existen algunas excepciones.
No alfa

En los organismos existen algunos aminoácidos no α. En estas estructuras, el grupo amina se desplazó más lejos del extremo ácido carboxílico de la molécula de aminoácido. Por lo tanto, un aminoácido β tiene el grupo amina unido al segundo carbono, y un aminoácido γ lo tiene en el tercero. Los ejemplos incluyen β-alanina , GABA y ácido δ- aminolevulínico .

β-alanina: un aminoácido producido por la aspartato 1-descarboxilasa y un precursor de la coenzima A y los péptidos carnosina y anserina .

Ácido γ-aminobutírico (GABA): un neurotransmisor en animales.

Ácido δ-aminolevulínico: un intermedio en la biosíntesis de tetrapirrol ( hemo , clorofila , cobalamina , etc. ).

Ácido 4-aminobenzoico (PABA): un intermedio en la biosíntesis de folato



La razón por la que los α-aminoácidos se utilizan en proteínas se ha relacionado con su frecuencia en meteoritos y experimentos prebióticos. Una especulación inicial sobre las propiedades deletéreas de los β-aminoácidos en términos de estructura secundaria resultó ser incorrecta.
D-aminoácidos
Algunos aminoácidos contienen la quiralidad absoluta opuesta, sustancias químicas que no están disponibles en la maquinaria normal de traducción / transcripción de ribosomas. La mayoría de las paredes de las células bacterianas están formadas por peptidoglicano , un polímero compuesto de aminoazúcares entrecruzados con oligopéptidos cortos unidos entre sí. El oligopéptido no se sintetiza ribosómicamente y contiene varias peculiaridades que incluyen D-aminoácidos , generalmente D-alanina y D-glutamato. Una peculiaridad adicional es que el primero es racemizado por enzimas de unión a PLP (codificadas por alr o el homólogo dadX ), mientras que el segundo es racemizado por una enzima independiente de cofactor ( murI ). Algunas variantes están presentes en Thermotoga spp. La D-lisina está presente y en ciertas bacterias resistentes a la vancomicina está presente la D-serina ( gen vanT ).
En los animales, algunos D-aminoácidos son neurotransmisores.
Sin hidrógeno en el carbono α
Todos los aminoácidos proteinogénicos tienen al menos un hidrógeno en el carbono α. La glicina tiene dos hidrógenos y todos los demás tienen un hidrógeno y una cadena lateral. La sustitución del hidrógeno restante por un sustituyente más grande, como un grupo metilo, distorsiona la estructura de la proteína.
En algunos hongos , el ácido α-aminoisobutírico se produce como precursor de los péptidos, algunos de los cuales exhiben propiedades antibióticas. Este compuesto es similar a la alanina, pero posee un grupo metilo adicional en el carbono α en lugar de un hidrógeno. Por tanto, es aquiral. Otro compuesto similar a la alanina sin un α-hidrógeno es la deshidroalanina , que posee una cadena lateral de metileno. Es uno de varios deshidroaminoácidos naturales .

Alanina

ácido aminoisobutírico

deshidroalanina



Estereocentros gemelos de aminoácidos
Un subconjunto de L-α-aminoácidos es ambiguo en cuanto a cuál de los dos extremos es el carbono α. En las proteínas, un residuo de cisteína puede formar un enlace disulfuro con otro residuo de cisteína, reticulando así la proteína. Dos cisteínas reticuladas forman una molécula de cistina . La cisteína y la metionina se producen generalmente por sulfurilación directa, pero en algunas especies pueden producirse por transulfuración , donde la homoserina o serina activada se fusiona con una cisteína u homocisteína que forma cistationina . Un compuesto similar es lantionina , que puede verse como dos moléculas de alanina unidas mediante un enlace tioéter y se encuentra en varios organismos. De manera similar, el ácido djenkólico , una toxina vegetal de los frijoles jengkol , está compuesto por dos cisteínas conectadas por un grupo metileno. El ácido diaminopimélico se usa como puente en el peptidoglicano y se usa como precursor de la lisina (a través de su descarboxilación).

cistina
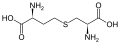
cistationina

lantionina
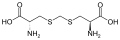
Ácido djenkólico
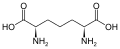
Ácido diaminopimélico

Aminoácidos prebióticos y bioquímicas alternativas
En meteoritos y en experimentos prebióticos (por ejemplo, el experimento de Miller-Urey ) se encuentran muchos más aminoácidos que los veinte aminoácidos estándar, varios de los cuales en concentraciones más altas que los estándar: se ha conjeturado que si surgiera la vida basada en aminoácidos en paralelo en otras partes del universo, no más del 75% de los aminoácidos serían comunes. La anomalía más notable es la falta de ácido aminobutírico.
| Proporción de aminoácidos en relación con la glicina (%) | ||
|---|---|---|
| Molécula | Descarga eléctrica | Meteorito de Murchinson |
| glicina | 100 | 100 |
| Alanina | 180 | 36 |
| ácido α-amino-n-butírico | 61 | 19 |
| norvalina | 14 | 14 |
| valina | 4.4 | |
| norleucina | 1.4 | |
| leucina | 2.6 | |
| isoleucina | 1.1 | |
| aloisoleucina | 1.2 | |
| t-leucina | <0,005 | |
| ácido α-amino-n-heptanoico | 0,3 | |
| prolina | 0,3 | 22 |
| ácido pipecólico | 0,01 | 11 |
| ácido α, β-diaminopropiónico | 1,5 | |
| ácido α, γ-diaminobutírico | 7,6 | |
| ornitina | <0.01 | |
| lisina | <0.01 | |
| ácido aspártico | 7.7 | 13 |
| ácido glutamico | 1,7 | 20 |
| serina | 1.1 | |
| treonina | 0,2 | |
| alotreonina | 0,2 | |
| metionina | 0,1 | |
| homocisteína | 0,5 | |
| homoserina | 0,5 | |
| β-alanina | 4.3 | 10 |
| ácido β-amino-n-butírico | 0,1 | 5 |
| ácido β-aminoisobutírico | 0,5 | 7 |
| ácido γ-aminobutírico | 0,5 | 7 |
| ácido α-aminoisobutírico | 7 | 33 |
| isovalina | 1 | 11 |
| sarcosina | 12,5 | 7 |
| N-etilglicina | 6,8 | 6 |
| N-propil glicina | 0,5 | |
| N-isopropil glicina | 0,5 | |
| N-metil alanina | 3.4 | 3 |
| N-etil alanina | <0.05 | |
| N-metil β-alanina | 1.0 | |
| N-etil β-alanina | <0.05 | |
| isoserina | 1.2 | |
| ácido α-hidroxi-γ-aminobutírico | 17 |
Cadena lateral recta
El código genético se ha descrito como un accidente congelado y las razones por las que solo hay un aminoácido estándar con una cadena lineal (alanina) podrían ser simplemente la redundancia con valina, leucina e isoleucina. Sin embargo, se informa que los aminoácidos de cadena lineal forman hélices alfa mucho más estables.

Glicina (cadena lateral de hidrógeno)
Alanina (cadena lateral de metilo)

Homoalanina o ácido α-aminobutírico (cadena lateral de etilo)

Norvalina ( cadena lateral n -propilo)

Norleucina ( cadena lateral de n- butilo)

Homonorleucina ( cadena lateral n- pentilo) (se muestra ácido heptanoico)




Calcógeno
La serina, homoserina , O-metil-homoserina y O-etil-homoserina poseen una cadena lateral de hidroximetilo, hidroxietilo, O-metil-hidroximetilo y O-metil-hidroxietilo. Mientras que la cisteína, homocisteína , metionina y etionina poseen los equivalentes de tiol. Los equivalentes de selenol son selenocisteína, selenohomocisteína, selenometionina y selenoetionina. Los aminoácidos con la siguiente capa de calcógeno también se encuentran en la naturaleza: varias especies como Aspergillus fumigatus, Aspergillus terreus y Penicillium chrysogenum en ausencia de azufre son capaces de producir e incorporar a las proteínas telurocisteína y telurometionina .
La hidroxiglicina, un aminoácido con una cadena lateral de hidroxilo, es muy inestable.
Código genético ampliado
Roles
En las células, especialmente en las autótrofas, se encuentran varios aminoácidos no proteinogénicos como intermediarios metabólicos. Sin embargo, a pesar de la flexibilidad catalítica de las enzimas de unión a PLP, muchos aminoácidos se sintetizan como cetoácidos ( por ejemplo, 4-metil-2-oxopentanoato a leucina) y se aminan en el último paso, manteniendo así el número de aminoácidos no proteinogénicos. intermedios bastante bajos.
La ornitina y la citrulina se encuentran en el ciclo de la urea , parte del catabolismo de los aminoácidos (ver más abajo).
Además del metabolismo primario, varios aminoácidos no proteinogénicos son precursores de la producción final en el metabolismo secundario para producir pequeños compuestos o péptidos no ribosomales (como algunas toxinas ).
Incorporado postraduccionalmente en proteína
A pesar de no estar codificados por el código genético como aminoácidos proteinogénicos, algunos aminoácidos no estándar se encuentran en las proteínas. Estos se forman mediante la modificación postraduccional de las cadenas laterales de los aminoácidos estándar presentes en la proteína diana. Estas modificaciones suelen ser esenciales para la función o regulación de una proteína; por ejemplo, en el gamma-carboxiglutamato, la carboxilación del glutamato permite una mejor unión de los cationes de calcio , y en la hidroxiprolina, la hidroxilación de la prolina es fundamental para mantener los tejidos conectivos . Otro ejemplo es la formación de hipusina en el factor de iniciación de la traducción EIF5A , mediante la modificación de un residuo de lisina. Tales modificaciones también pueden determinar la localización de la proteína, por ejemplo, la adición de grupos hidrófobos largos puede hacer que una proteína se una a una membrana fosfolipídica .

Ácido carboxiglutámico. Mientras que el ácido glutámico posee un grupo γ-carboxilo, el ácido carboxiglutámico posee dos.

Hidroxiprolina. Este iminoácido se diferencia de la prolina debido a un grupo hidroxilo en el carbono 4.
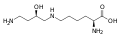
Hipusina . Este aminoácido se obtiene agregando al grupo ε-amino de una lisina un resto 4-aminobutilo (obtenido a partir de espermidina )


-Pyroglutamic_acid_Structural_Formulae.png)
Existe alguna evidencia preliminar de que el ácido aminomalónico puede estar presente, posiblemente por incorporación errónea, en las proteínas.
Análogos tóxicos
Varios aminoácidos no proteinogénicos son tóxicos debido a su capacidad para imitar ciertas propiedades de los aminoácidos proteinogénicos, como la tialisina . Algunos aminoácidos no proteinogénicos son neurotóxicos al imitar los aminoácidos utilizados como neurotransmisores (es decir, no para la biosíntesis de proteínas), por ejemplo , ácido quiscualico , canavanina o ácido azetidina-2-carboxílico . La cefalosporina C tiene una cadena principal de ácido α-aminoadípico (homoglutamato) que está amidada con un resto de cefalosporina. La penicilamina es un aminoácido terapéutico cuyo mecanismo de acción se desconoce.

Tialisina

Ácido quiscualico

Canavanine

ácido azetidina-2-carboxílico

Cefalosporina C

Penicilamina

-Azetidine-2-carboxylate.png)

Las cianotoxinas de origen natural también pueden incluir aminoácidos no proteinogénicos. La microcistina y la nodularina , por ejemplo, se derivan ambas de ADDA , un β-aminoácido.
No aminoácidos
La taurina es un ácido amino sulfónico y no un ácido amino carboxílico, sin embargo, ocasionalmente se considera como tal, ya que las cantidades necesarias para suprimir el auxótrofo en ciertos organismos (por ejemplo, gatos) son más cercanas a las de los "aminoácidos esenciales" (auxotrofia de aminoácidos). que de las vitaminas (cofactor auxotrofia).
Los osmolitos, la sarcosina y la glicina betaína se derivan de los aminoácidos, pero tienen una amina secundaria y una cuaternaria respectivamente.